










Slate-throated Redstart Myioborus miniatus Scientific name definitions
- LC Least Concern
- Names (28)
- Subspecies (12)
William D. Harrod and Ronald L. Mumme
Version: 2.0 — Published July 29, 2022
Revision Notes
Revision Notes
Sign in to see your badges
Species names in all available languages
| Language | Common name |
|---|---|
| Catalan | bosquerola pissarrosa |
| Dutch | Meniezanger |
| English | Slate-throated Redstart |
| English (United States) | Slate-throated Redstart |
| French | Paruline ardoisée |
| French (France) | Paruline ardoisée |
| German | Larvenwaldsänger |
| Japanese | ベニイタダキアメリカムシクイ |
| Norwegian | skiferhvitstjert |
| Polish | pleszówka ciemnogłowa |
| Portuguese (Brazil) | mariquita-cinza |
| Portuguese (Portugal) | Mariquita-cinzenta |
| Russian | Черногорлая канделита |
| Serbian | Crvenokapa američka crvenrepka |
| Slovak | horárik bridlicový |
| Spanish | Candelita Plomiza |
| Spanish (Argentina) | Arañero Garganta Negra |
| Spanish (Costa Rica) | Candelita Pechinegra |
| Spanish (Ecuador) | Candelita Goliplomiza |
| Spanish (Honduras) | Chipe Pavito Garganta Ceniza |
| Spanish (Mexico) | Pavito Alas Negras |
| Spanish (Panama) | Candelita Gargantiplomiza |
| Spanish (Peru) | Candelita de Garganta Plomiza |
| Spanish (Spain) | Candelita plomiza |
| Spanish (Venezuela) | Candelita Gargantipizarra |
| Swedish | skiffervitstjärt |
| Turkish | Kül Rengi Ötleğen |
| Ukrainian | Чернітка чорногорла |
Revision Notes
William D. Harrod and Ronald L. Mumme revised this account. Arnau Bonan Barfull curated the media.
Myioborus miniatus (Swainson, 1827)
PROTONYM:
Setophaga miniata
Swainson, 1827. The Philosophical Magazine, New Series n.s., 1, p.368.
TYPE LOCALITY:
woods of Valladolid [= Morelia], Michoacan.
SOURCE:
Avibase, 2023
Definitions
- MYIOBORUS
- miniata / miniatus
The Key to Scientific Names
Legend Overview
UPPERCASE: current genus
Uppercase first letter: generic synonym
● and ● See: generic homonyms
lowercase: species and subspecies
●: early names, variants, misspellings
‡: extinct
†: type species
Gr.: ancient Greek
L.: Latin
<: derived from
syn: synonym of
/: separates historical and modern geographic names
ex: based on
TL: type locality
OD: original diagnosis (genus) or original description (species)
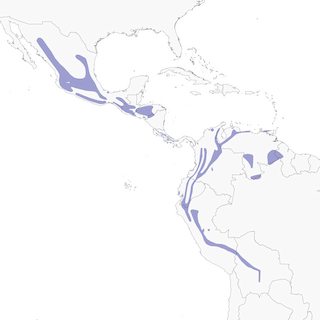
- Year-round
- Migration
- Breeding
- Non-Breeding
Distribution of the Slate-throated Redstart















