

Species names in all available languages
| Language | Common name |
|---|---|
| Catalan | becard gorja-rosat |
| Dutch | Grote Bekarde |
| English | Rose-throated Becard |
| English (United States) | Rose-throated Becard |
| French | Bécarde à gorge rose |
| French (France) | Bécarde à gorge rose |
| German | Rosenkehlbekarde |
| Japanese | バラノドカザリドリモドキ |
| Norwegian | rosenstrupebekard |
| Polish | bekardzik purpurowogardły |
| Russian | Розовогорлый бекард |
| Serbian | Ružogrli bekard |
| Slovak | tityra mexická |
| Spanish | Anambé Degollado |
| Spanish (Costa Rica) | Cabezón Plomizo |
| Spanish (Honduras) | Cabezón Garganta Rosada |
| Spanish (Mexico) | Cabezón Degollado |
| Spanish (Panama) | Cabezón Plomizo |
| Spanish (Spain) | Anambé degollado |
| Swedish | rosastrupig bekard |
| Turkish | Gülrengi Boğazlı Bekard |
| Ukrainian | Бекард великий |
Pachyramphus aglaiae (de Lafresnaye, 1839)
Definitions
- PACHYRAMPHUS
- aglaiae
The Key to Scientific Names
Legend Overview
Rose-throated Becard Pachyramphus aglaiae Scientific name definitions
Version: 1.0 — Published March 4, 2020
Text last updated February 14, 2014
Distribution
Introduction
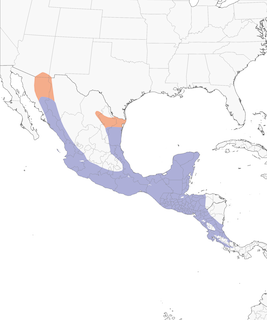
Breeding range: Abundance in its range varies locally and regionally from common and fairly common to rare. Species appears to be most prevalent in tropical dry forest and riparian woodland in semi-arid or arid regions and least so in tropical wet forest areas. Breeding range is restricted to Middle America, extending from the Mexican borderlands of southeastern Arizona and southeastern Texas, south through Costa Rica, and perhaps to Chiriquí Province of western Panama (Ridgely and Gwynne 1989; eBird.org database [see below]). The range splits in the north around the arid, interior Mexican Plateau, with prongs following the slopes of the Sierra Madre Oriental and Sierra Madre Occidental in northern Mexico, and extending into wooded habitats in the sierran foothills and along some drainages into the adjoining lowlands.
The status of the species in some regions within its range or at its southern limits remains uncertain or unknown, notably in the large area of the Caribbean lowla3nds (see main distribution map) of northeastern Honduras (Gracias a Dios) and adjoining northeastern Nicaragua (Atlantíco Norte), in the eastern Caribbean slope and lowlands of Costa Rica, and in western Panama (Chiriqui Province at least). The first region may be blank in our range map simply because it is relatively remote and seldom visited, both historically and now. In the last two regions, this becard was described as "a rare visitor, probably a migrant", found only from November to March, in Costa Rica (Stiles and Skutch 1989: 293, Garrigues 2007), and as "Apparently a casual winter visitant to lower highlands of western Chiriquí" during the northern winter months, perhaps as "migrants" from the north (Ridgely and Gwynne 1989: 318). These speculations about migrants are without support. No specimens of migratory populations of P. a. albiventris are known south of the breeding ranges of this subspecies. Also, a growing number of reports in both regions, especially in southeastern Costa Rica and westernmost Panama, suggest that the species may be regular year round and more widely distributed than previously thought. In northwestern Costa Rica at least, birds have been reported through the end of May (breeding season), and from early August on (NMNH [see acronyms beyond], Jiménez specimens; eBird.org reports, 2005-13). On the Pacific side of Costa Rica near the Panama border, some observers visiting in April (early breeding season) have reported them (eBird data). In Chiriquí Province, one observer described the becard that he watched as a dark form (D. Rogers, 22 February 2013; eBird.org), not a pale northern type. It should be noted that most observers visit these regions during the drier winter months, not during the local rainy seasons. However, the status of this becard in Caribbean and southeastern (Pacific) Costa Rica and the status and limits of distribution in western Panama remain to be clarified.
Figure 5 provides a more detailed view of distribution color-coded to show the approximate ranges of the eight subspecies (see above) currently recognized (Snow 1979). The locations of specimen records displayed in the map come from published sources of Ridgway (1907), Hellmayr (1929), and Webster (1963), and from specimens determined to subspecies rank in the holdings of the National Museum of Natural History (NMNH), Field Museum of Natural History, Museum of Comparative Zoology, American Museum of Natural History, and California Academy of Sciences (CAS), which occurred within or near regions expected to harbor the subspecies (Webster 1963). Ridgway’s (1907) and Hellmayr’s (1929) locational data of the nominate subspecies included localities that we now know to fall within the range of the northeastern form, P. a. gravis, so the records used here from those sources are mapped to reflect current understanding. Also, I supplemented the specimen records with information supplied in the literature on the results of local collecting expeditions to Mexico (Sutton and Burleigh 1939, Brodkorb 1943, Wetmore 1943, Blake 1950, Lowery and Dalquest 1951, Warner and Mengel 1951, Zimmerman and Harry 1951, Tashian 1952, Davis 1953, Selander and Giller 1959, Urban 1959, Short 1974, Binford 1989) and on critical reviews of regional avifaunas (Dickey and van Rossem 1938, Paynter 1955, Russell 1964, Smithe 1966, Monroe 1968, Oberholser 1974, Russell and Monson 1998 (locational data provided by S. M. Russell, in litt.). As a result of the last work, the distribution of the species in Sonora is better understood than in any other region of similar size in Middle America. I also have used some field reports posted to the eBird database at the Cornell Lab of Ornithology, especially those based on observations in southeastern Costa Rica (potential breeding season reports) and nearby western Panama, as noted earlier. Zones of intergradation between subspecies need additional work. Populations in the Motagua River valley in Guatemala and at Copán in Honduras have been controversial (Dickey and van Rossem 1938, Webster 1963, Monroe 1968) and deserve further attention. Three subspecies, P. a. sumichrasti, P. a. hypophaeus, and P. a. latirostris, meet in this region. The range limits of the three subspecies in the area, and their genetic contribution to the controversial populations, are not yet resolved. Other contact areas worthy of attention are in Guerrero and Morelos, Mexico, between P. a. albiventris and P. a. aglaiae; in southern Quintana Roo, Mexico, and northern Belize between P. a. yucatanensis and P. a. hypophaeus; in southern Campeche and Tabasco, Mexico, between P. a. yucatanensis and P. a. sumichrasti; and from the northern border of Veracruz to central Veracruz, Mexico, between P. a. gravis, P. a. sumichrasti, and P. a. aglaiae. In the last area, the designated type locality of P. a. aglaiae at Jalapa [= Xalapa-Enríquez], Veracruz (Nelson 1897, Webster 1963: 393; Figure 5), appears to be in this zone of intergradation. The status of P. a. aglaiae as a separate subspecies is an open question as well (Webster 1963), while the others are relatively distinct. Apart from subspecies, genetic and morphological variation across the range of the species deserves to be investigated using modern methods.
The dynamics of becard populations in the Mexican borderland regions of Texas (mainly Hidalgo County) and Arizona (mainly Santa Cruz County), where they are at the northern edge of their range, may not be typical of populations away from range limits. In both regions, birds always have been rare to uncommon, and populations local and probably very small, and restricted to riparian habitats in the lower Rio Grande Valley in Texas (Lockwood and Freeman 2004, Brush 2005), and in similar habitat along sections of the Santa Cruz River, especially along Sonoita Creek near Patagonia, in Arizona (Monson and Phillips 1981, Corman 2005). Irregular occurrences of breeding birds at most sites and frequent nest failures may be expected in such regions. Appendix 1 summarizes the observational history of becards reported in the seasonal reports for both regions since 1970 and by the Texas Bird Records Committee (TBRC). Only some of these reports have been documented and reviewed by the TBRC, but most other reports probably also are dependable, given that the species is distinctive and attracts interest. The status of this species has changed in both regions during the period of these reports. During the 1970s, becards were relatively prevalent locally in Texas, notably in Santa Ana National Wildlife Refuge and Anzalduas County Park. Evidently, the species was sufficiently regular then to be under-reported in the seasonal reports covering this area (Appendix 1 ). Brush (2005, 2008) reviewed this early history and noted that becards nested successfully in the region from the 1940s through the 1970s. Between 1987-2011, 74% of 50 accepted occurrences (TBRC) were in Hidalgo Co., 14% in Cameron County, and 8% in Starr County. Evidently the earliest record is a specimen from Brownsville, Cameron County, 30 October 1891 (Oberholser 1974); the first reported nesting was in Cameron County in April, 1943 (Davis 1945). Successful nests and some failed attempts were documented in Texas from the 1940s to the late 1970s, followed by an apparent breeding hiatus until sporadic nestings occurred during the 1990s through the early 2000s that were unsuccessful (Gehlbach 1987, Brush 2005, 2008; Appendix 1 ). The most recent successful nests were at Santa Ana N.W.R and at Anzalduas County Park, Hidalgo County, in 1999 (Brush 2000). Also, one or two becards have begun appearing in restoration habitat in Estero Llano Grande State Park, Weslaco, and have overwintered there beginning in 2008–09.
In Arizona, the earliest record of this species was a young male taken in the Huachuca Mountains, Cochise County, 20 June 1888 (Price 1888, Phillips 1949, Webster 1963); two later reports also are from there (Taylor 1995, designated as accidental there). Later historical nesting records in that state outside of the expected range of the becard are from Guadalupe Canyon (Levy 1958) and from the Chiricahua Mountains (Gibbs and Gibbs 1956, but regarded as suspect by Monson and Phillips 1981), both in Cochise County, along Arivaca Creek, Pima County (Corman 2005), and in upper Aravaipa Canyon, Graham County (Monson and Phillips 1981). A single report of a bird regarded as "accidental" comes from the San Pedro River watershed just north of Fairbank, Cochise County, on 2 June 1989 (Krueper 1999; D.J. Krueper in litt.). During the 1970s, becards along Sonoita Creek near Patagonia were reported as "numerous" at times, while numbers were low in other years (Appendix 1 ). Monson and Phillips (1981) thought that the species might be "pushing northwards", but it was regular, at least through the late 1980s, only at one locality on Sonoita Creek. No reports of becards were posted for the period from 1988 through 1997 on Sonoita Creek (perhaps a case of under-reporting early during this time) until in 1998 when becards were noted as again present (Appendix 1 ). Another hiatus of reports ensued until the early 2000s when the seasonal editor for passerines in the region remarked that "numbers of this scarce species in Arizona have noticeably declined during the past decade" (Rosenberg and Stevenson 2002; Appendix 1 ).
So both in Texas and in Arizona, becards were more regular through the 1970s, and perhaps into the 1980s, even though they usually were found at one or two localities, and numbers varied from year to year. In both regions, birds were mostly absent from the few familiar areas during the 1990s, and now are sporadic and rarely even attempt to breed, or too often attempted nestings fail. Often in the last 20 years or more, only one or two individuals have been present for varying periods of times, with reports from every month of the year (see below).
Nonbreeding range and migration: The wintering range of becards that withdraw from the northernmost portions of their breeding range in Sonora, Mexico, and in adjoining Arizona, is unknown. No long distance movements are known from specimen evidence of the northern subspecies, contra speculations by Ridgely and Gwynne (1989) and Mobley (2004). Possibly, northern P. a. albiventris withdraws into the range of resident birds immediately to the south. Some individuals overwinter around Alamos, Sonora, whereas most birds are absent during the nonbreeding season (Russell 2000; S.M. Russell in litt.). A male was found far to the north of its wintering range at Arroyo Santo Domingo in Sonora, and beginning in 1983 (Russell and Monson 1988), birds have been present in winter on at least six occasions in Santa Cruz and Pima counties, Arizona (Appendix 1 ). From 1970–1989, becards occurred mainly from May into July, and only casually in fall or early winter. During 1990–2012, reports were concentrated during the breeding season, May–July, but individuals were found irregularly in all other months (Appendix 1 ). Thus, evidence for migration best applies to populations in southeastern Arizona and Sonora, where northward movement in Sonora proceeds gradually as individuals arrive beginning in mid-March in the south around Alamos and in early May in the north. Fall movement commences in August and continues into early October (Russell and Monson 1998).
The status of becards in the Lower Rio Grande Valley of Texas (P. a. gravis) varies annually and seasonally, but current information suggests that the population in the region (including northern Tamaulipas) is at best only partially migratory (contra Howell and Webb 1995). Historical reports are sparse, but I. Davis (Bent 1942) reported what he believed to be a single, immature male as present from October 1937 (initially with a female) into early December 1938 in Cameron County. Oberholser (1974) wrote that species was "rare, local, and irregular" from early October to late March in Hidalgo and Cameron counties. During the period 1970–1989, becard occurrences (individuals were registered for each month during a span of known presence) exhibited a bimodal pattern through the year, with a May–June peak during the breeding period and a strong December-February peak in winter. From 1990–2012, occurrences (n = 89) were spread across all months with a single peak during the winter (39%) (Appendix 1 ). Thus, currently the species appears to nest only sporadically and locally in Texas (Brush 2005) and are more frequent winter visitors. Occurrence and seasonality of Rose-throated Becards in the adjoining Mexican states bordering Texas (northern Tamaulipas and northern Nuevo Leon) are largely unknown (see below).
No direct information is available on dispersal in Rose-throated Becards. However, it is likely that populations in the borderlands of southeastern Arizona and southeastern Texas are maintained by dispersal and recruitment from nearby populations in Mexico. Little is known about these populations, but perhaps wooded river valleys in Mexico near the border provide corridors to riparian habitats in these two states, mainly the Santa Cruz River and a main tributary, Sonoita Creek, south of Tucson, and along the Rio Grande on the Texas side of the river. Flesch (2008) surveyed the birds in northern Sonora, including habitats in the Río Concepcion watershed that approach Arizona’s Santa Cruz River near the border towns of Nogales. Rose-throated Becards were present at four localities in this watershed, including along the headwater tributaries Río Cocospera/Río Bambuto nearest the border, but they were considered "rare" wherever they were found. The urban development around the cities of Nogales, Sonora, and Nogales, Arizona, may be filter barriers to dispersal farther north. The Río Gila watershed, which includes the Mexican portion of the Santa Cruz River, also offers a source of becards for dispersal to the north into Arizona. The species was encountered there along the Santa Cruz River in Sonora, but again was regarded as "rare". The nearest known populations of becards to the habitat in Texas are in Monterrey, Nuevo León (Sutton and Burleigh 1939, Brush 2005), and farther south in Linares (Sutton and Pettingill 1943, Eaton and Edwards 1947). Brush (in litt.) reported that the species is common along stream courses within and near the Sierra Madre Oriental. They also are resident along the Río Corona near Ciudad Victoria about 200 km south-southwest of Santa Ana National Wildlife Refuge (Brush 2005), but habitat may be scarce in northern Tamaulipas because of extensive agricultural development on the coastal plain. The exceptions may be Río San Fernando, which has woodland in its riparian zone, and Río San Juan, which flows out of the Sierra Madre Oriental into the Rio Grande River about 55 km to the west of favored habitats in Texas.
The unpredictable appearances and disappearances of becards along the lower Rio Grande in Texas implicates post-breeding movements and wandering from Mexico. Evidence of post-breeding dispersal in the species is weak, but such movements probably accounts for the presence of birds in Texas and Arizona in fall and winter. Webster (1963) suggested that some individual becards of this species, especially young males, tend to wander upslope into the highlands (or to more northerly localities) in fall and winter, thus accounting for their appearance at unusually high elevations above 1500 m in temperate localities.
Elsewhere, Forcey (2002) reported a "strong annual movement" into the riparian corridors where they breed at over 1500 m in the mountains of Oaxaca. The scarcity of becards in these mountain corridors in the nonbreeding season suggests elevational movements, probably mostly downslope to lower valleys, but a few birds occur at very high elevations (below). Farther south in the range of the species, postbreeding movements may cause birds to appear where they seldom are found. In Costa Rica, the species has occurred as an apparent altitudinal "vagrant" on the Caribbean slope in the upper Peñas Blancas valley near Monteverde (Fogden 1993).
Extralimital occurrences: Extralimital occurrences of wandering birds outside the Lower Rio Grande Valley in Texas are from Chambers County (Oberholser 1974), Jeff Davis County (Runnels 1975, Peterson and Zimmer 1998), Jim Wells County (Rappole and Blacklock 1985), Kenedy County (Haynie 1993), and Aransas County (Lockwood 2002). In Arizona, reports from upper Aravaipa Canyon and Huachuca Mountains, and perhaps from Guadalupe Canyon, are unusual occurrences well away from the known, more regular breeding sites in the upper Santa Cruz watershed. One record for New Mexico is from upper Guadalupe Canyon, Hidalgo County (9 June 1979; Witzeman et al. 1979). Unverified reports come from Big Bend National Park, Texas (Wauer 1996). Farther south in the becard's range where birds are resident, post-breeding movement may cause birds to appear where they are seldom known.
Rose-throated Becard is restricted to mainland North America. No extralimital occurrences beyond its continental range are known.
Historical Changes to the Distribution
Rose-throated Becard formerly was more frequent than it is now as a breeding species in the border regions of southeast Texas and south central Arizona. Habitat loss in neighboring areas of Mexico that cut off avenues of recruitment in its peripheral range in the United States may be an issue. Sutton (1949) also recorded habitat loss from agricultural land clearing near McAllen, Texas, as responsible for the loss of an historical breeding site in a lakeshore woodland. Otherwise, little is known on this topic elsewhere in the range of the species.
- Year-round
- Migration
- Breeding
- Non-Breeding

eBird Status and Trends















