










Red-crested Cardinal Paroaria coronata Scientific name definitions
- LC Least Concern
- Names (29)
- Monotypic
Amanda Linn, Kevin J. Burns, and Casey H. Richart
Version: 1.0 — Published March 4, 2020
Text last updated July 10, 2015
Text last updated July 10, 2015
Sign in to see your badges
Species names in all available languages
| Language | Common name |
|---|---|
| Catalan | sit cardenal crestat |
| Chinese | 紅冠唐納雀 |
| Dutch | Roodkuifkardinaal |
| English | Red-crested Cardinal |
| English (United States) | Red-crested Cardinal |
| French | Paroare huppé |
| French (France) | Paroare huppé |
| German | Rotschopftangare |
| Japanese | コウカンチョウ |
| Norwegian | gråkardinal |
| Polish | kardynałek czubaty |
| Portuguese (Brazil) | cardeal |
| Portuguese (Portugal) | Cardeal-sul-americano |
| Russian | Краснохохлая танагра-кардинал |
| Serbian | Crvenokruni kardinal |
| Slovak | kardinálka sivá |
| Slovenian | Mehiški kardinal |
| Spanish | Cardenilla Crestada |
| Spanish (Argentina) | Cardenal Copete Rojo |
| Spanish (Chile) | Cardenal |
| Spanish (Ecuador) | Cardenal Crestirrojo |
| Spanish (Paraguay) | Cardenal |
| Spanish (Peru) | Cardenal de Cresta Roja |
| Spanish (Puerto Rico) | Cardenal Copetirrojo |
| Spanish (Spain) | Cardenilla crestada |
| Spanish (Uruguay) | Cardenal Copete Rojo |
| Swedish | rödtofsad kardinaltangara |
| Turkish | Kırmızı Tepeli Kardinal |
| Ukrainian | Пароарія чубата |
Paroaria coronata (Miller, 1776)
PROTONYM:
Loxia coronata
Miller, 1776. Icones Animalium: Various subjects of natural history, wherein are delineated birds, animals and many curious plants, with the parts of fructification of each plant, all of which are drawn and coloured from nature pt1, pl.2.
TYPE LOCALITY:
no type locality [reference not verified] ; Rio Grande do Sul, Brazil, designated by Pinto, 1944, Cat. Aves Brasil (Publ. Dept. Zool., Sao Paulo), pt. 2, p. 602.
SOURCE:
Avibase, 2023
Definitions
- PAROARIA
- coronata
The Key to Scientific Names
Legend Overview
UPPERCASE: current genus
Uppercase first letter: generic synonym
● and ● See: generic homonyms
lowercase: species and subspecies
●: early names, variants, misspellings
‡: extinct
†: type species
Gr.: ancient Greek
L.: Latin
<: derived from
syn: synonym of
/: separates historical and modern geographic names
ex: based on
TL: type locality
OD: original diagnosis (genus) or original description (species)
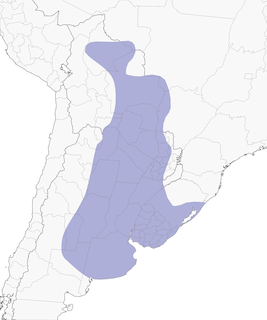
- Year-round
- Migration
- Breeding
- Non-Breeding
Distribution of the Red-crested Cardinal















