












Peruvian Sheartail Thaumastura cora Scientific name definitions
- LC Least Concern
- Names (21)
- Monotypic
Christopher J. Clark
Version: 1.0 — Published March 4, 2020
Text last updated August 23, 2013
Text last updated August 23, 2013
Sign in to see your badges
Species names in all available languages
| Language | Common name |
|---|---|
| Catalan | colibrí de Cora |
| Czech | kalypta břitvochvostá |
| Dutch | Peruaanse Schaarstaartkolibrie |
| English | Peruvian Sheartail |
| English (United States) | Peruvian Sheartail |
| French | Colibri cora |
| French (France) | Colibri cora |
| German | Corakolibri |
| Japanese | ホソフタオハチドリ |
| Norwegian | pinsettkolibri |
| Polish | brzytwosterek |
| Russian | Перуанский искрохвост |
| Slovak | čmeľovec britvochvostý |
| Spanish | Colibrí Cora |
| Spanish (Chile) | Picaflor de Cora |
| Spanish (Ecuador) | Colifina Peruana |
| Spanish (Peru) | Colibrí de Cora |
| Spanish (Spain) | Colibrí cora |
| Swedish | långstjärtad skogsjuvel |
| Turkish | Tel Kuyruklu Çölyıldızı |
| Ukrainian | Колібрі перуанський |
Thaumastura cora (Lesson & Garnot, 1827)
PROTONYM:
Orthorhynchus Cora
Lesson & Garnot, 1827. Voyage autour du Monde exécuté par ordre du Roi, sur la Corvette de Sa Majesté, La Coquille, pendant les années 1822, 1823, 1824 et 1825 (etc.). Zoologie 1, p.682, pl.31 fig.4.
TYPE LOCALITY:
Between Callao and Lima, Peru.
SOURCE:
Avibase, 2023
Definitions
- THAUMASTURA
- cora
- Cora
The Key to Scientific Names
Legend Overview
UPPERCASE: current genus
Uppercase first letter: generic synonym
● and ● See: generic homonyms
lowercase: species and subspecies
●: early names, variants, misspellings
‡: extinct
†: type species
Gr.: ancient Greek
L.: Latin
<: derived from
syn: synonym of
/: separates historical and modern geographic names
ex: based on
TL: type locality
OD: original diagnosis (genus) or original description (species)
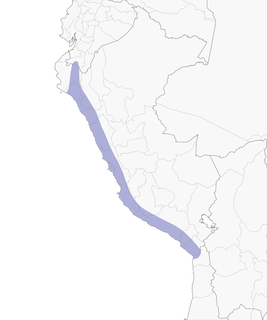
- Year-round
- Migration
- Breeding
- Non-Breeding
Distribution of the Peruvian Sheartail















