
















Olivaceous Woodcreeper Sittasomus griseicapillus Scientific name definitions
- Names (30)
- Subspecies (15)
Michael A. Patten
Version: 1.0 — Published March 4, 2020
Text last updated January 19, 2011
Text last updated January 19, 2011
Sign in to see your badges
Species names in all available languages
| Language | Common name |
|---|---|
| Catalan | grimpa-soques olivaci meridional |
| Dutch | Grijze Muisspecht |
| English | Olivaceous Woodcreeper |
| English (United States) | Olivaceous Woodcreeper |
| French | Grimpar fauvette |
| French (France) | Grimpar fauvette |
| German | Dünnschnabel-Baumsteiger |
| Japanese | ヒメオニキバシリ |
| Norwegian | gråhodetreløper |
| Polish | kowal |
| Portuguese (Brazil) | arapaçu-verde |
| Portuguese (Portugal) | Arapaçu-verde |
| Russian | Поползневый древолаз |
| Serbian | Maslinasta puzavica |
| Slovak | kôrolezec sivohlavý |
| Spanish | Trepatroncos Oliváceo |
| Spanish (Argentina) | Tarefero |
| Spanish (Costa Rica) | Trepadorcito Aceitunado |
| Spanish (Ecuador) | Trepatroncos Oliváceo |
| Spanish (Honduras) | Trepatroncos Oliváceo |
| Spanish (Mexico) | Trepatroncos Cabeza Gris |
| Spanish (Panama) | Trepatroncos Oliváceo |
| Spanish (Paraguay) | Guirí |
| Spanish (Peru) | Trepador Oliváceo |
| Spanish (Spain) | Trepatroncos oliváceo |
| Spanish (Uruguay) | Tarefero |
| Spanish (Venezuela) | Trepador Verdón |
| Swedish | gråhuvad trädklättrare |
| Turkish | Zeytuni Tırmaşık |
| Ukrainian | Дереволаз оливковий |
Sittasomus griseicapillus (Vieillot, 1818)
PROTONYM:
Dendrocopus griseicapillus
Vieillot, 1818. Nouveau dictionnaire d'histoire naturelle, appliquée aux arts, à l'agriculture, à l'économie rurale et domestique, à la médecine, etc. Nouvelle édition 26, p.119.
TYPE LOCALITY:
Paraguay; based on 'trepadore palido y roxo' of Azara; restricted to Concepcion del Paraguay by Hellmayr, 1925.
SOURCE:
Avibase, 2023
Definitions
- SITTASOMUS
- griseicapilla / griseicapillus
The Key to Scientific Names
Legend Overview
UPPERCASE: current genus
Uppercase first letter: generic synonym
● and ● See: generic homonyms
lowercase: species and subspecies
●: early names, variants, misspellings
‡: extinct
†: type species
Gr.: ancient Greek
L.: Latin
<: derived from
syn: synonym of
/: separates historical and modern geographic names
ex: based on
TL: type locality
OD: original diagnosis (genus) or original description (species)
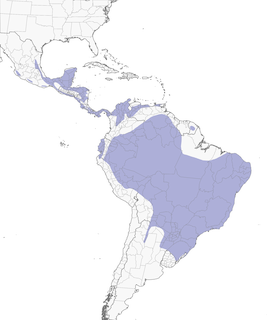
- Year-round
- Migration
- Breeding
- Non-Breeding
Distribution of the Olivaceous Woodcreeper















