











Black Swift Cypseloides niger Scientific name definitions
- VU Vulnerable
- Names (30)
- Subspecies (3)
Carolyn Gunn, Peter E. Lowther, Charles T. Collins, Jason Beason, Kim Potter, and M. Webb
Version: 3.0 — Published February 3, 2023
Revision Notes
Revision Notes
Sign in to see your badges
Species names in all available languages
| Language | Common name |
|---|---|
| Catalan | falciot fosc |
| Czech | rorýs černý |
| Dutch | Zwarte Gierzwaluw |
| English | Black Swift |
| English (United States) | Black Swift |
| French | Martinet sombre |
| French (France) | Martinet sombre |
| German | Schwarzsegler |
| Haitian Creole (Haiti) | Chiksòl |
| Icelandic | Svartsvölungur |
| Japanese | クロムジアマツバメ |
| Norwegian | svartseiler |
| Polish | cierniosternik czarny |
| Portuguese (Brazil) | taperuçu-escuro |
| Russian | Американский стриж |
| Serbian | Američka crna čiopa |
| Slovak | sadziar čierny |
| Spanish | Vencejo Negro |
| Spanish (Costa Rica) | Vencejo Negro |
| Spanish (Cuba) | Vencejo negro |
| Spanish (Dominican Republic) | Vencejo Negro |
| Spanish (Honduras) | Vencejo Negro |
| Spanish (Mexico) | Vencejo Negro |
| Spanish (Panama) | Vencejo Negro |
| Spanish (Puerto Rico) | Vencejo Negro |
| Spanish (Spain) | Vencejo negro |
| Spanish (Venezuela) | Vencejo Negro |
| Swedish | svart seglare |
| Turkish | Karaca Ebabil |
| Ukrainian | Свіфт західний |
Revision Notes
Carolyn Gunn revised the account, with contributions from Peter Pyle on the Plumages, Molts, and Structure page. Arnau Bonan Barfull curated the media.
Cypseloides niger (Gmelin, 1789)
PROTONYM:
Hirundo nigra
Gmelin, 1789. Systema Naturae. Editio decima tertia, aucta, reformata. Cura Jo. Frid. Gmelin (etc.). Tomus I. Pars II 1 pt2, p.1025.
TYPE LOCALITY:
Hispaniola.
SOURCE:
Avibase, 2023
Definitions
- CYPSELOIDES
- cypseloides
- niger
The Key to Scientific Names
Legend Overview
UPPERCASE: current genus
Uppercase first letter: generic synonym
● and ● See: generic homonyms
lowercase: species and subspecies
●: early names, variants, misspellings
‡: extinct
†: type species
Gr.: ancient Greek
L.: Latin
<: derived from
syn: synonym of
/: separates historical and modern geographic names
ex: based on
TL: type locality
OD: original diagnosis (genus) or original description (species)
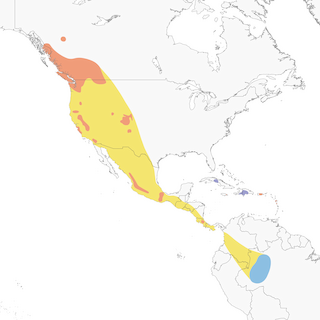
- Year-round
- Migration
- Breeding
- Non-Breeding
Distribution of the Black Swift















