












Bay-headed Tanager Tangara gyrola Scientific name definitions
- LC Least Concern
- Names (25)
- Subspecies (9)
Cyndarienne Mireles and Kevin J. Burns
Version: 1.0 — Published March 4, 2020
Text last updated September 27, 2010
Text last updated September 27, 2010
Sign in to see your badges
Species names in all available languages
| Language | Common name |
|---|---|
| Catalan | tàngara cap-rogenca |
| Dutch | Okerkaptangare |
| English | Bay-headed Tanager |
| English (United States) | Bay-headed Tanager |
| French | Calliste rouverdin |
| French (France) | Calliste rouverdin |
| German | Rötelkopftangare |
| Japanese | チャガシラフウキンチョウ |
| Norwegian | brunhodetanagar |
| Polish | tangarka rudogłowa |
| Portuguese (Brazil) | saíra-de-cabeça-castanha |
| Portuguese (Portugal) | Saíra-de-cabeça-castanha |
| Russian | Рыжеголовая танагра |
| Serbian | Zelena tangara |
| Slovak | tangara hnedohlavá |
| Spanish | Tangara Cabecibaya |
| Spanish (Costa Rica) | Tangara Cabecicastaña |
| Spanish (Ecuador) | Tangara Cabecibaya |
| Spanish (Panama) | Tangara Cabecibaya |
| Spanish (Peru) | Tangara de Cabeza Baya |
| Spanish (Spain) | Tangara cabecibaya |
| Spanish (Venezuela) | Tángara Cabeza de Lacre |
| Swedish | rosthuvad tangara |
| Turkish | Kızıl Başlı Tangara |
| Ukrainian | Гирола |
Tangara gyrola (Linnaeus, 1758)
PROTONYM:
Fringilla Gyrola
Linnaeus, 1758. Systema Naturæ per Regna Tria Naturæ, Secundum Classes, Ordines, Genera, Species, cum Characteribus, Differentiis, Synonymis, Locis. Tomus I. Editio decima, reformata 1, p.181.
TYPE LOCALITY:
Surinam.
SOURCE:
Avibase, 2023
Definitions
- TANGARA
- gyrola
- Gyrola
The Key to Scientific Names
Legend Overview
UPPERCASE: current genus
Uppercase first letter: generic synonym
● and ● See: generic homonyms
lowercase: species and subspecies
●: early names, variants, misspellings
‡: extinct
†: type species
Gr.: ancient Greek
L.: Latin
<: derived from
syn: synonym of
/: separates historical and modern geographic names
ex: based on
TL: type locality
OD: original diagnosis (genus) or original description (species)
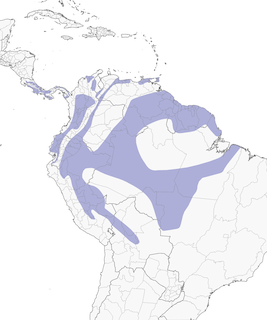
- Year-round
- Migration
- Breeding
- Non-Breeding
Distribution of the Bay-headed Tanager















